
久留米工業大学工学部教育創造工学科・教授・博士(理学)
主に昆虫を研究しています。
山地に住むクロヒカゲ(Lethe diana)というジャノメチョウ亜科の蝶は日本全土に広く分布し、京都では年三回成虫が出現します。幼虫は竹・笹類を食べ、成虫は樹液や動物の死体の汁を吸います。成虫の出現時期は、初夏世代が5-6月、盛夏世代が7-8月、秋世代が9-10月です。世代によって異なる気温条件下に置かれますが、季節型はないので、飛ぶためにはどの世代も同じ体温を維持する必要があると考えられます。
成虫の飛翔活動は午前と午後に一回ずつ盛んになります。この飛翔活動の盛んな時刻は初夏と秋には午前は遅い時間に、午後は早い時間にずれ、盛夏には逆に午前は早い時間に、午後は遅い時間にずれています。つまりほぼ同じ気温の時にさかんに飛んでいると考えられます。また、成虫は初夏には盛夏に比べ照度の高い場所を好み、日光浴をするなど、気温が低い時は日光の放射熱を利用していました。成虫の好む照度が季節的に変化するので、生息場所内の移動経路も季節によって位置が変わり、初夏は多少明るい場所を、盛夏は日陰の暗い場所を飛んでいました(Ide 2000a)。
成虫の調査地内の分布にも明るさは影響していました。成虫は雌雄とも餌資源である樹液が多い場所に集中分布していましたが、初夏と秋は明るい林縁部の狭い範囲に分布の中心ができ、盛夏には林内に空間的に連続して広がる暗い日陰に分布が移っていました(Ide 2000b)。そのため初夏には成虫の移動が抑制されましたが盛夏には広く移動し、個体群の空間構造は季節的に変化していました(Ide 2002a)。
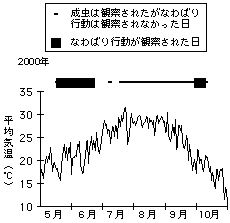
クロヒカゲの雄は日当りの良い木の梢に交尾のための縄張を作ります。縄張行動が見られるのは初夏と秋の午後だけで、盛夏には全く見られません(右図)(Ide 2002a)。成虫の体温を野外で実測したところ、縄張を作っていた個体の体温の範囲は23.8℃-33.6℃でした。蝶の体温は気温や日射に強く影響されることが知られています。これらの環境条件を終日測定したところ、初夏世代の雄が縄張行動を行っていたのは、体温を上記の範囲に保つのにちょうど良い温度環境が成立している時間帯だけでした。盛夏には午後に温度環境が好ましい範囲にまで下がってくるのは通常午後5時以降でした。つまり、配偶行動に季節変化が生じたのは、盛夏はほとんどの時間が暑すぎて縄張行動ができないためと考えられます(Ide 2002b)。
縄張行動が見られない盛夏に雄と雌がどのようにして出会っているのか調べたところ、主に午前中に雄が飛び回って雌を探索していることがわかりました。この午前中の雄による探索行動は他の季節にも見られ、初夏・秋世代の雄は午前中は飛び回って雌を探索し、午後には縄張を構えて雌を待伏せる、という具合に二つの行動を時間的に使い分けていました。クロヒカゲの羽化は朝に行われます。羽化したての未交尾雌は雄にとって価値が高く、従って朝の雌は平均的に価値が高いと考えられます。午後には羽化から時間がたち、残り寿命が減り、また既交尾率が上がることから、平均的な雌の価値は低くなります。従って、雌の価値が一日のうちに低下していくことになります。このような場合、後述する数理モデルが予測したように雌の価値が高い時には雄はコストが大きくても雌と出会いやすいように行動し、雌の価値が下がると雌と出会いにくくてもコストが小さくなるように行動して翌朝を待つことが有利になります。探索行動は雌を見つけやすい一方コストが大きく、縄張行動は雌と出会いにくいかわりに飛行コストなどは小さいと考えられますので、本種の雄の行動の変化はこの説明があてはまっていると思われます(Ide 2004b)。
多くの動物では、オスはたとえメスが拒否してもメスにしつこくつきまとって交尾しようとします。このようなオスのハラスメントがあると、メスの採餌時間や産卵時間が減ったり天敵に襲われる危険が増したりします。そのため、オスのハラスメントを避けるためのメスの行動が進化していることが予想されます。
蝶のベニシジミ(Lycaena phlaeas daimio)のメスを観察したところ、オスが近くを通りかかると翅を閉じました。また、メスが翅を閉じると、翅を開きっぱなしの場合よりもオスから求愛されることが減りました。以上の結果は、ベニシジミのメスの翅を閉じる行動にはオスの求愛を回避する機能があることを示しています。
この翅を閉じる行動は、主に既に一度交尾したメスで観察され、一度も交尾したことのないメスでは見られませんでした。ベニシジミのメスは、通常は生涯に一度しか交尾をしません。今後交尾をしないメスが行なうことから、この行動は間接的な配偶者選択(求愛行動の能力が高い雄を選ぶ)ではなくオスのしつこい求愛(ハラスメント)を回避するための行動と考えられます(Ide 2011)。
昆虫の配偶行動には雌雄それぞれにレック型(レック・群飛・配偶縄張など、雄が雌を待ち伏せ、雌が雄のいる場所を訪れる様式)と非レック型(雄が雌を動き回って探し、雌は雄が求愛して来るのを待つ様式)の二種類の行動様式があるとされています。同じ型の行動をとっている個体同士が交尾する場合、異性の同じ型の行動をとっている個体の割合が多いほど有利であり、他個体の行動によって繁殖成功率は変化します。クロヒカゲのようにレック型と非レック型を時刻によって使い分ける配偶戦略が進化するのはどんな場合かを明らかにするため、ゲームモデルを用いた解析を行いました。
モデルからは(1)レック型の雄のコストが少ない、(2)レック型の雌のコストが少ない、または(3)レック型の行動をとると雌雄が出会いやすい、という条件下でレック型の行動が進化することが予測されました。更に、雄にとっての雌の価値が一日のうちに減少する場合、雌の価値が高い時には雄はコストが大きくても雌と出会いやすい行動を採り、雌の価値が下がると雌と出会いにくくてもコストの小さい行動を採って翌朝を待つことが有利であり、コストの大きい行動から小さい行動へと配偶行動が切り替わる場合もあることが明らかになりました(Ide & Kondoh 2000)。また、雌雄間にゲーム理論的状況があることによって、雌雄間で配偶行動のタイミングを一致させる結果、ユスリカの蚊柱のような時間的に局限された配偶行動が進化することがあることが明らかになりました(Kondoh & Ide 2003)。

植食性昆虫にとって、一本の植物の中でも餌としての質には大きな変異があるので、植物上での摂食部位の選択は重要です。チュウゴクザサとカラムシを食草としている昆虫を対象に、食草上のどのような場所を選択しているのか、質の良い部分を利用できているのか、について調査しました。
チュウゴクザサ:本種の葉は数年の寿命がありますが、一度冬を越すと縁の部分が枯れて隈取ができます。このような葉は餌としての質が落ちるので食べるとよく成長できませんが、クロヒカゲの幼虫は隈取のない当年葉も隈取のある越冬葉もなぜか全く区別せずに利用していました(Ide 2003)。一方、同じチュウゴクザサの葉を巻いて巣を作り、巣を食べて生長するコチャバネセセリ(Thoressa varia)の幼虫は、当年葉だけを利用していました。隈取部分を切り取った越冬葉を与えると当年葉と同じように摂食したことから、隈取の有無を認識して葉の新旧を区別していることが分かりました(Ide 2004a)。
カラムシ:アカタテハ(Vanessa indica)の幼虫はカラムシの葉を折って巣を作り、巣を食べて成長します。巣を作る際に葉の付根の葉脈をかじって傷つけますが、この行動には葉が折れやすくなって巣を作りやすくなるという効果があります(Ide 2004c)。アカタテハの若齢幼虫は顎の発達がまだ不十分で葉脈を傷つけることができないため、曲げやすい芽の近くの葉を利用していました。しかし、成長して葉脈を傷つけられるようになると、芽の近くの葉は避けてもう少し古くて硬い葉を選択するようになりました(Ide 2009)。やはりカラムシを餌とする蛾のフクラスズメ(Arcte coerulea)の幼虫は、しばしば大発生して葉を食い尽くすことが知られています。フクラスズメは芽の付近の葉を避けるので、カラムシは葉を食い尽くされても芽から素早く葉を展開し光合成を再開することができます。カラムシはフクラスズメの大発生に対して、芽に防御物質を貯めて対抗していると考えられます(Ide 2006a)。
2015年7月27日
御意見、御質問は下のメールアドレスまでお寄せ下さい。
idejy [at] kurume-it.ac.jp